Statistics
Statistics
241 human active and 13 inactive phosphatases in total;
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update:
11 Mar, 2019
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update:
11 Mar, 2019
Top
PFKFB1
| Gene Name | PFKFB1 (QuickGO) | (A quick tutorial to explore the interctive visulaization) |
| Synonyms | PFKFB1, F6PK, PFRX | |
| Protein Name | PFKFB1 |
|
| Alternative Name(s) |
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 1;6PF-2-K/Fru-2,6-P2ase 1;PFK/FBPase 1;6PF-2-K/Fru-2,6-P2ase liver isozyme;6-phosphofructo-2-kinase;2.7.1.105;Fructose-2,6-bisphosphatase;3.1.3.46; | |
| EntrezGene ID | 5207 (Comparitive Toxicogenomics) | |
| UniProt AC (Human) | P16118 (protein sequence) | |
| Enzyme Class | EC 2.7.1.105 3.1.3.46 (BRENDA | |
| Molecular Weight | 54681 Dalton | |
| Protein Length | 471 amino acids (AA) | |
| Genome Browsers | NCBI | ENSG00000158571 (Ensembl) | UCSC | |
| Crosslinking annotations | Query our ID-mapping table | |
| Orthologues | Quest For Orthologues (QFO) | GeneTree | |
| Classification | Superfamily: histidine phosphatase (HP) --> Family: PFKFB | Historic class: Phosphoglycerate mutase | CATH ID: 3.40.50.1240 | SCOP Fold: HP | |
| Phosphatase activity | active | Catalytic signature motif: unknown | |
| Phosphorylation Network | Visualize | |
| Domain organization, Expression, Diseases | (show / hide) |
| Protein Domain | InterPro | Pfam | SMART | 3D Structure | PDB | DrugPort | ModBase | SwissModel |
| Polypeptide organization (Pfam) | No Pfam domain found | ||
| Catalytic Site | Mechanism and Catalytic Site Atlas | ||
| Gene Expression | Gene Expression Atlas | Diseases | OMIM | ClinVar |
| Protein-protein interactions | STRING | IntAct | MINT | BioGRID | ||
| Phosphosites | Phospho.ELM | PhosphoSitePlus | ||
| Localization, Function, Catalytic activity and Sequence | (show / hide) |
| Localization (UniProt annotation) | |||
| N/A | |||
| Function (UniProt annotation) | |||
| Synthesis and degradation of fructose 2,6-bisphosphate | |||
| Catalytic Activity (UniProt annotation) | |||
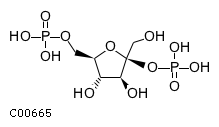
| Beta-D-fructose 2,6-bisphosphate + H(2)O = D-fructose 6-phosphate + phosphate ATP + D-fructose 6-phosphate = ADP + beta-D-fructose 2,6-bisphosphate | |||
| Protein Sequence | |||
| MSPEMGELTQTRLQKIWIPHSSGSSRLQRRRGSSIPQFTNSPTMVIMVGLPARGKTYISTKLTRYLNWIGTPTKVFNLGQ YRREAVSYKNYEFFLPDNMEALQIRKQCALAALKDVHNYLSHEEGHVAVFDATNTTRERRSLILQFAKEHGYKVFFIESI CNDPGIIAENIRQVKLGSPDYIDCDREKVLEDFLKRIECYEVNYQPLDEELDSHLSYIKIFDVGTRYMVNRVQDHIQSRT VYYLMNIHVTPRSIYLCRHGESELNIRGRIGGDSGLSVRGKQYAYALANFIQSQGISSLKVWTSHMKRTIQTAEALGVPY EQWKALNEIDAGVCEEMTYEEIQEHYPEEFALRDQDKYRYRYPKGESYEDLVQRLEPVIMELERQENVLVICHQAVMRCL LAYFLDKSSDELPYLKCPLHTVLKLTPVAYGCKVESIYLNVEAVNTHREKPENVDITREPEEALDTVPAHY |
| Motif information from Eukaryotic Linear Motif atlas (ELM) | (show / hide) |
| ELM Motif | Motif Instance | Description | Motif Regex |
| NA | NA | NA | NA |
| Gene Ontology (P: Process; F: Function and C: Component terms) | (show / hide) | |
| GO:0003873 | F:6-phosphofructo-2-kinase activity |
| GO:0004331 | F:fructose-2,6-bisphosphate 2-phosphatase activity |
| GO:0005524 | F:ATP binding |
| GO:0005829 | C:cytosol |
| GO:0006000 | P:fructose metabolic process |
| GO:0006003 | P:fructose 2,6-bisphosphate metabolic process |
| GO:0006094 | P:gluconeogenesis |
| GO:0006096 | P:glycolytic process |
| GO:0019900 | F:kinase binding |
| GO:0031100 | P:animal organ regeneration |
| GO:0032868 | P:response to insulin |
| GO:0033133 | P:positive regulation of glucokinase activity |
| GO:0033762 | P:response to glucagon |
| GO:0042594 | P:response to starvation |
| GO:0042802 | F:identical protein binding |
| GO:0043540 | C:6-phosphofructo-2-kinase/fructose-2,6-biphosphatase complex |
| GO:0045820 | P:negative regulation of glycolytic process |
| GO:0045821 | P:positive regulation of glycolytic process |
| GO:0051384 | P:response to glucocorticoid |
| GO:0051591 | P:response to cAMP |
| GO:0070095 | F:fructose-6-phosphate binding |