241 human active and 13 inactive phosphatases in total;
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update: 11 Mar, 2019
Cytoplasm Cell membrane Membrane raft Cell projection, axon Cytoplasm, cytoskeleton Note=Localizes at nerve terminalsin brain and at bundles of microtubules surrounding the nucleus inthe elongating spermatids corresponding to the manchette (Bysimilarity) Translocates from the cytoplasm to membrane rufflesin a RAC1-dependent manner (PubMed:11084340)
Function (UniProt annotation)
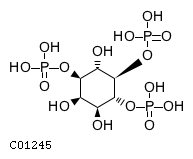
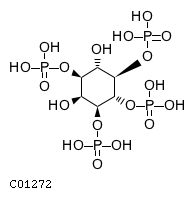
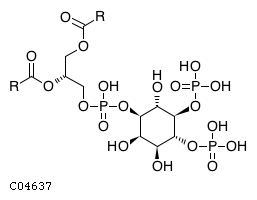
Inositol 5-phosphatase which may be involved in distinctmembrane trafficking and signal transduction pathways May mediatethe inhibitory effect of Rac1 on endocytosis
At the plasma membrane, subsequent phosphorylation of phosphatidylinositol 4-phosphate (PI4P) produces phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) and phosphatidylinositol 3,4,5-trisphosphate (PI(3,4,5)P3) while the actions of various other kinases and phosphatases produces phosphatidylinositol 3-phosphate (PI3P), phosphatidylinositol 5-phosphate (PI5P), phosphatidylinositol 3,4-bisphosphate (PI(3,4)P2), and phosphatidylinositol 3,5-bisphosphate (PI(3,5)P2) (Zhang et al. 1997, Gurung et al. 2003, Guo et al. 1999, Vanhaesebroeck et al. 1997, Tolias et al. 1998, Schaletzky et al. 2003, Kim et al. 2002, Clarke et al. 2010). Many of the phosphatidylinositol phosphatases that act at the plasma membrane belong to the myotubularin family. Enzymatically inactive myotubularin family members can heterodimerize with catalytically active mytotubularins to regulate their stability, activity and/or substrate specificity (Berger et al. 2006, Zou et al. 2012)
Clathrin-mediated endocytosis (CME) is one of a number of process that control the uptake of material from the plasma membrane, and leads to the formation of clathrin-coated vesicles (Pearse et al, 1975; reviewed in Robinson, 2015; McMahon and Boucrot, 2011; Kirchhausen et al, 2014). CME contributes to signal transduction by regulating the cell surface expression and signaling of receptor tyrosine kinases (RTKs) and G-protein coupled receptors (GPCRs). Most RTKs exhibit a robust increase in internalization rate after binding specific ligands; however, some RTKs may also exhibit significant ligand-independent internalization (reviewed in Goh and Sorkin, 2013). CME controls RTK and GPCR signaling by organizing signaling both within the plasma membrane and on endosomes (reviewed in Eichel et al, 2016; Garay et al, 2015; Vieira et al, 1996; Sorkin and von Zastrow, 2014; Di Fiori and von Zastrow, 2014; Barbieri et al, 2016). CME also contributes to the uptake of material such as metabolites, hormones and other proteins from the extracellular space, and regulates membrane composition by recycling membrane components and/or targeting them for degradation. Clathrin-mediated endocytosis involves initiation of clathrin-coated pit (CCP) formation, cargo selection, coat assembly and stabilization, membrane scission and vesicle uncoating. Although for simplicity in this pathway, the steps leading to a mature CCP are represented in a linear and temporally distinct fashion, the formation of a clathrin-coated vesicle is a highly heterogeneous process and clear temporal boundaries between these processes may not exist (see for instance Taylor et al, 2011; Antonescu et al, 2011; reviewed in Kirchhausen et al, 2014). Cargo selection in particular is a critical aspect of the formation of a mature and stable CCP, and many of the proteins involved in the initiation and maturation of a CCP contribute to cargo selection and are themselves stabilized upon incorporation of cargo into the nascent vesicle (reviewed in Kirchhausen et al, 2014; McMahon and Boucrot, 2011).Although the clathrin triskelion was identified early as a major component of the coated vesicles, clathrin does not bind directly to membranes or to the endocytosed cargo. Vesicle formation instead relies on many proteins and adaptors that can bind the plasma membrane and interact with cargo molecules. Cargo selection depends on the recognition of endocytic signals in cytoplasmic tails of the cargo proteins by adaptors that interact with components of the vesicle's inner coat. The classic adaptor for clathrin-coated vesicles is the tetrameric AP-2 complex, which along with clathrin was identified early as a major component of the coat. Some cargo indeed bind directly to AP-2, but subsequent work has revealed a large family of proteins collectively known as CLASPs (clathrin- associated sorting proteins) that mediate the recruitment of diverse cargo into the emerging clathrin-coated vesicles (reviewed in Traub and Bonifacino, 2013). Many of these CLASP proteins themselves interact with AP-2 and clathrin, coordinating cargo recruitment with coat formation (Schmid et al, 2006; Edeling et al, 2006; reviewed in Traub and Bonifacino, 2013; Kirchhausen et al, 2014). Initiation of CCP formation is also influenced by lipid composition, regulated by clathrin-associated phosphatases and kinases (reviewed in Picas et al, 2016). The plasma membrane is enriched in PI(4,5)P2. Many of the proteins involved in initiating clathrin-coated pit formation bind to PI(4,5)P2 and induce membrane curvature through their BAR domains (reviewed in McMahon and Boucrot, 2011; Daumke et al, 2014). Epsin also contributes to early membrane curvature through its Epsin N-terminal homology (ENTH) domain, which promotes membrane curvature by inserting into the lipid bilayer (Ford et al, 2002). Following initiation, some CCPs progress to formation of vesicles, while others undergo disassembly at the cell surface without producing vesicles (Ehrlich et al, 2004; Loerke et al, 2009; Loerke et al, 2011; Aguet et al, 2013; Taylor et al, 2011). The assembly and stabilization of nascent CCPs is regulated by several proteins and lipids (Mettlen et al, 2009; Antonescu et al, 2011).Maturation of the emerging clathrin-coated vesicle is accompanied by further changes in the lipid composition of the membrane and increased membrane curvature, promoted by the recruitment of N-BAR domain containing proteins (reviewed in Daumke et al, 2014; Ferguson and De Camilli, 2012; Picas et al, 2016). Some N-BAR domain containing proteins also contribute to the recruitment of the large GTPase dynamin, which is responsible for scission of the mature vesicle from the plasma membrane (Koh et al, 2007; Lundmark and Carlsson, 2003; Soulet et al, 2005; David et al, 1996; Owen et al, 1998; Shupliakov et al, 1997; Taylor et al, 2011; Ferguson et al, 2009; Aguet et al, 2013; Posor et al, 2013; Chappie et al, 2010; Shnyrova et al, 2013; reviewed in Mettlen et al, 2009; Daumke et al, 2014). After vesicle scission, the clathrin coat is dissociated from the new vesicle by the ATPase HSPA8 (also known as HSC70) and its DNAJ cofactor auxilin, priming the vesicle for fusion with a subsequent endocytic compartment and releasing clathrin for reuse (reviewed in McMahon and Boucrot, 2011; Sousa and Laufer, 2015)

 Statistics
Statistics