Statistics
Statistics
241 human active and 13 inactive phosphatases in total;
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update:
11 Mar, 2019
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update:
11 Mar, 2019
Top
LPIN2
| Gene Name | LPIN2 (QuickGO) | (A quick tutorial to explore the interctive visulaization) |
| Synonyms | LPIN2, KIAA0249 | |
| Protein Name | LPIN2 |
|
| Alternative Name(s) |
Phosphatidate phosphatase LPIN2;3.1.3.4;Lipin-2; | |
| EntrezGene ID | 9663 (Comparitive Toxicogenomics) | |
| UniProt AC (Human) | Q92539 (protein sequence) | |
| Enzyme Class | EC 3.1.3.4 (BRENDA | |
| Molecular Weight | 99399 Dalton | |
| Protein Length | 896 amino acids (AA) | |
| Genome Browsers | NCBI | ENSG00000101577 (Ensembl) | UCSC | |
| Crosslinking annotations | Query our ID-mapping table | |
| Orthologues | Quest For Orthologues (QFO) | GeneTree | |
| Classification | Superfamily: HAD --> Family: Lipin | Historic class: Lipin family | CATH ID: 3.40.50.1000 | SCOP Fold: HAD | |
| Phosphatase activity | active | Catalytic signature motif: unknown | |
| Phosphorylation Network | Visualize | |
| Domain organization, Expression, Diseases | (show / hide) |
| Protein Domain | InterPro | Pfam | SMART | 3D Structure | PDB | DrugPort | ModBase | SwissModel |
| Polypeptide organization (Pfam) | No Pfam domain found | ||
| Catalytic Site | Mechanism and Catalytic Site Atlas | ||
| Gene Expression | Gene Expression Atlas | Diseases | OMIM | COSMIC | ClinVar |
| Protein-protein interactions | STRING | IntAct | MINT | BioGRID | ||
| Phosphosites | Phospho.ELM | PhosphoSitePlus | ||
| Localization, Function, Catalytic activity and Sequence | (show / hide) |
| Localization (UniProt annotation) | |||
| Nucleus Cytoplasm, cytosol Endoplasmic reticulum membrane Note=Translocates to endoplasmic reticulum membrane withincreasing levels of oleate | |||
| Function (UniProt annotation) | |||
| Plays important roles in controlling the metabolism offatty acids at different levels Acts as a magnesium-dependentphosphatidate phosphatase enzyme which catalyzes the conversion ofphosphatidic acid to diacylglycerol during triglyceride,phosphatidylcholine and phosphatidylethanolamine biosynthesis inthe reticulum endoplasmic membrane Acts also as a nucleartranscriptional coactivator for PPARGC1A to modulate lipidmetabolism (By similarity) | |||
| Catalytic Activity (UniProt annotation) | |||
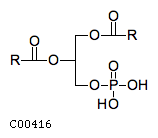
| A 1,2-diacylglycerol 3-phosphate + H(2)O = a1,2-diacyl-sn-glycerol + phosphate | |||
| Protein Sequence | |||
| MNYVGQLAGQVIVTVKELYKGINQATLSGCIDVIVVQQQDGSYQCSPFHVRFGKLGVLRSKEKVIDIEINGSAVDLHMKL GDNGEAFFVEETEEEYEKLPAYLATSPIPTEDQFFKDIDTPLVKSGGDETPSQSSDISHVLETETIFTPSSVKKKKRRRK KYKQDSKKEEQAASAAAEDTCDVGVSSDDDKGAQAARGSSNASLKEEECKEPLLFHSGDHYPLSDGDWSPLETTYPQTAC PKSDSELEVKPAESLLRSESHMEWTWGGFPESTKVSKRERSDHHPRTATITPSENTHFRVIPSEDNLISEVEKDASMEDT VCTIVKPKPRALGTQMSDPTSVAELLEPPLESTQISSMLDADHLPNAALAEAPSESKPAAKVDSPSKKKGVHKRSQHQGP DDIYLDDLKGLEPEVAALYFPKSESEPGSRQWPESDTLSGSQSPQSVGSAAADSGTECLSDSAMDLPDVTLSLCGGLSEN GEISKEKFMEHIITYHEFAENPGLIDNPNLVIRIYNRYYNWALAAPMILSLQVFQKSLPKATVESWVKDKMPKKSGRWWF WRKRESMTKQLPESKEGKSEAPPASDLPSSSKEPAGARPAENDSSSDEGSQELEESITVDPIPTEPLSHGSTTSYKKSLR LSSDQIAKLKLHDGPNDVVFSITTQYQGTCRCAGTIYLWNWNDKIIISDIDGTITKSDALGQILPQLGKDWTHQGIAKLY HSINENGYKFLYCSARAIGMADMTRGYLHWVNDKGTILPRGPLMLSPSSLFSAFHREVIEKKPEKFKIECLNDIKNLFAP SKQPFYAAFGNRPNDVYAYTQVGVPDCRIFTVNPKGELIQERTKGNKSSYHRLSELVEHVFPLLSKEQNSAFPCPEFSSF CYWRDPIPEVDLDDLS |
| Motif information from Eukaryotic Linear Motif atlas (ELM) | (show / hide) |
| ELM Motif | Motif Instance | Description | Motif Regex |
| NA | NA | NA | NA |
| Gene Ontology (P: Process; F: Function and C: Component terms) | (show / hide) | |
| GO:0003713 | F:transcription coactivator activity |
| GO:0005634 | C:nucleus |
| GO:0005789 | C:endoplasmic reticulum membrane |
| GO:0005829 | C:cytosol |
| GO:0006351 | P:transcription |
| GO:0006629 | DNA-templated |
| GO:0006646 | P:lipid metabolic process |
| GO:0006656 | P:phosphatidylethanolamine biosynthetic process |
| GO:0008195 | P:phosphatidylcholine biosynthetic process |
| GO:0009062 | F:phosphatidate phosphatase activity |
| GO:0019432 | P:fatty acid catabolic process |
| GO:0045944 | P:triglyceride biosynthetic process |