
 Statistics
Statistics
241 human active and 13 inactive phosphatases in total;
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update:
11 Mar, 2019
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update:
11 Mar, 2019
CHEBI:17345
| Name | guanosine 5'-monophosphate | 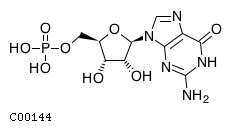 Download: mol | sdf |
| Synonyms | 05'-gmp; 5'-guanylic acid; 5'-o-phosphonatoguanosine; Gmp; Guanosine 5'-monophosphate; Guanosine 5'-phosphate; Guanosine monophosphate; Guanylic acid; | |
| Definition | A purine ribonucleoside 5'-monophosphate having guanine as the nucleobase. | |
| Molecular Weight (Exact mass) | 363.2206 (363.058) | |
| Molecular Formula | C10H14N5O8P | |
| SMILES | Nc1nc2n(cnc2c(=O)[nH]1)[C@@H]1O[C@H](COP(O)(O)=O)[C@@H](O)[C@H]1O | |
| InChI | InChI=1S/C10H14N5O8P/c11-10-13-7-4(8(18)14-10)12-2-15(7)9-6(17)5(16)3(23-9)1-22-24(19,20)21/h2-3,5-6,9,16-17H,1H2,(H2,19,20,21)(H3,11,13,14,18)/t3-,5-,6-,9-/m1/s1 | |
| InChI Key | RQFCJASXJCIDSX-UUOKFMHZSA-N | |
| Crosslinking annotations | KEGG:C00144 | 3DMET:B01172 | CAS:85-32-5 | ChEBI:17345 | ChEMBL:CHEMBL283807 | KNApSAcK:C00019635 | NIKKAJI:J10.615A | PDB-CCD:5GP | PDB-CCD:G | PubChem:3444 | |

| Pathway ID | Pathway Name | Pathway Description (KEGG) |
| map00230 | Purine metabolism | NA |
| map01060 | Biosynthesis of plant secondary metabolites | NA |
| map01065 | Biosynthesis of alkaloids derived from histidine and purine | NA |
| map01100 | Metabolic pathways | NA |
| map01523 | Antifolate resistance | Since the 1940s, antifolates have played a pivotal role in drug treatment of malignant, microbial, parasitic and chronic inflammatory diseases. The molecular basis of the anti-proliferative activity of antifolates relies on inhibition of key enzymes in folate metabolism, which results in disruption of purine and thymidylate biosynthesis, inhibition of DNA replication and cell death. The anti-inflammatory properties of antifolate have been most strongly related to its ability to block the release of pro-inflammatory cytokines such as tumour necrosis factor (TNF)-alpha or interleukin (IL)-1beta. Cells may develop resistance to an antifolate drug by virtue of impaired drug transport into cells, augmented drug export, impaired activation of antifolates through polyglutamylation, augmented hydrolysis of antifolate polyglutamates, increased expression and mutation of target enzymes, and the augmentation of cellular tetrahydrofolate-cofactor pools in cells. |
| map04022 | cGMP-PKG signaling pathway | Cyclic GMP (cGMP) is the intracellular second messenger that mediates the action of nitric oxide (NO) and natriuretic peptides (NPs), regulating a broad array of physiologic processes. The elevated intracellular cGMP level exerts its physiological action through two forms of cGMP-dependent protein kinase (PKG), cGMP-regulated phosphodiesterases (PDE2, PDE3) and cGMP-gated cation channels, among which PKGs might be the primary mediator. PKG1 isoform-specific activation of established substrates leads to reduction of cytosolic calcium concentration and/or decrease in the sensitivity of myofilaments to Ca2+ (Ca2+-desensitization), resulting in smooth muscle relaxation. In cardiac myocyte, PKG directly phosphorylates a member of the transient potential receptor canonical channel family, TRPC6, suppressing this nonselective ion channel's Ca2+ conductance, G-alpha-q agonist-induced NFAT activation, and myocyte hypertrophic responses. PKG also opens mitochondrial ATP-sensitive K+ (mitoKATP) channels and subsequent release of ROS triggers cardioprotection. |
| map04740 | Olfactory transduction | Within the compact cilia of the olfactory receptor neurons (ORNs) a cascade of enzymatic activity transduces the binding of an odorant molecule to a receptor into an electrical signal that can be transmitted to the brain. Odorant molecules bind to a receptor protein (R) coupled to an olfactory specific Gs-protein (G) and activate a type III adenylyl cyclase (AC), increasing intracellular cAMP levels. cAMP targets an olfactory-specific cyclic-nucleotide gated ion channel (CNG), allowing cations, particularly Na and Ca, to flow down their electrochemical gradients into the cell, depolarizing the ORN. Furthermore, the Ca entering the cell is able to activate a Ca-activated Cl channel, which would allow Cl to flow out of the cell, thus further increasing the depolarization. Elevated intracellular Ca causes adaptation by at least two different molecular steps: inhibition of the activity of adenylyl cyclase via CAMKII-dependent phosphorylation and down-regulation of the affinity of the CNG channel to cAMP. |
| map04742 | Taste transduction | Five basic tastes are recognized by humans and most other animals - bitter, sweet, sour, salty and umami. In vertebrates, taste stimuli are detected by taste receptor cells (TRCs). At least three distinct cell types are found in mammalian taste buds : type I cells, type II cells, and type III cells. Type I cells express epithelial sodium channel (ENaC) and are considered to be the major mediator of perception of low salt. In type II cells, transduction of bitter, sweet and umami is mediated by a canonical PLC-beta/IP3-signaling cascade, which culminates in the opening of the TRPM5 ion channel. This produces a depolarization that may allow CALMH1 channels to open and release ATP, which serves as a neurotransmitter to activate closely associated nerve afferents expressing P2X2, P2X3 receptors and adjacent type III cells expressing P2Y4 receptors. Type II taste cells also secrete acetylcholine (ACh) that appears to stimulate muscarinic receptors, specifically M3, on the same or neighboring Type II cells. This muscarinic feedback augments taste-evoked release of ATP. In type III cells, sour taste is initiated when protons enter through apically located proton-selective ion channels: polycystic kidney disease 2-like 1 protein (PKD2L1) and polycystic kidney disease 1-like 3 protein (PKD1L3) channels. Weak acids may also activate sour cells by penetrating the cell membrane and leading to closure of resting K+ channels and membrane depolarization. Further, voltage-gated Ca2+ channels are activated and release vesicular serotonin (5-HT), norepinephrine (NE) and gamma-aminobutyric acid (GABA). 5-HT and GABA provide negative paracrine feedback onto receptor cells by activating 5-HT1A and GABAA, GABAB receptors, respectively. 5-HT also functions as a transmitter between presynaptic cells and the sensory afferent. |
| map04744 | Phototransduction | Phototransduction is a biochemical process by which the photoreceptor cells generate electrical signals in response to captured photons. The vertebrate cascade starts with the absorption of photons by the photoreceptive pigments, the rhodopsins, which consist of a membrane embedded chromophore, 11-cis-retinal, and a G-protein-coupled receptor, opsin. The photon isomerizes 11-cis-retinal to all-trans-retinal which induces a structural change that activates the opsin. This triggers hydrolysis of cGMP by activating a transducinphosphodiesterase 6 (PDE6) cascade, which results in closure of the cGMP-gated cation channels (CNG) in the plasma membrane and membrane hyperpolarization. The hyperpolarization of the membrane potential of the photoreceptor cell modulates the release of neurotransmitters to downstream cells. Recovery from light involves the deactivation of the light- activated intermediates: photolyzed rhodopsin is phosphorylated by rhodopsin kinase (RK) and subsequently capped off by arrestin; GTP-binding transducin alpha subunit deactivates through a process that is stimulated by RGS9. |