241 human active and 13 inactive phosphatases in total;
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last scientific update: 11 Mar, 2019
Bifunctional enzyme (PubMed:12574510) The C-terminaldomain has epoxide hydrolase activity and acts on epoxides (alkeneoxides, oxiranes) and arene oxides (PubMed:12869654,PubMed:12574510, PubMed:22798687) Plays a role in xenobioticmetabolism by degrading potentially toxic epoxides (Bysimilarity) Also determines steady-state levels of physiologicalmediators (PubMed:12869654, PubMed:12574510, PubMed:22798687) TheN-terminal domain has lipid phosphatase activity, with the highestactivity towards threo-9,10-phosphonooxy-hydroxy-octadecanoicacid, followed by erythro-9,10-phosphonooxy-hydroxy-octadecanoicacid, 12-phosphonooxy-octadec-9Z-enoic acid and 12-phosphonooxy-octadec-9E-enoic acid (PubMed:12574510)
Catalytic Activity (UniProt annotation)
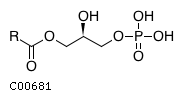
An epoxide + H(2)O = a glycol (9S,10S)-10-hydroxy-9-(phosphonooxy)octadecanoate + H(2)O = (9S,10S)-9,10-dihydroxyoctadecanoate + phosphate
Peroxisomes are essential organelles that play a key role in redox signalling and lipid homeostasis. They contribute to many crucial metabolic processes such as fatty acid oxidation, biosynthesis of ether lipids and free radical detoxification. The biogenesis of peroxisomes starts with the early peroxins PEX3, PEX16 and PEX19 and proceeds via several steps. The import of membrane proteins into peroxisomes needs PEX19 for recognition, targeting and insertion via docking at PEX3. Matrix proteins in the cytosol are recognized by peroxisomal targeting signals (PTS) and transported to the docking complex at the peroxisomal membrane. Peroxisomes' deficiencies lead to severe and often fatal inherited peroxisomal disorders (PD). PDs are usually classified in two groups. The first group is disorders of peroxisome biogenesis which include Zellweger syndrome, and the second group is single peroxisomal enzyme deficiencies.
The epoxidation of arachidonic acid by cytochrome P450s (CYPs) results in the formation of unique bioactive lipid mediators termed epoxyeicosatrienoic acids (EETs). Each double bond has been shown to be susceptible to oxidation, resulting in 5,6-EET, 8,9-EET, 11,12-EET, and 14,15-EET. The majority of the EET biological activities are diminished by the hydrolysis to the corresponding dihydroxyeicosatrienoic acids (DHET) (Capdevila et al. 2000, Buczynski et al. 2009, Vance & Vance 2008)
Maresins 1 and 2 (MaR1 and MaR2) are derived through the action of lipoxygenase 12 on the ω-3 fatty acid docosahexaenoic acid (DHA). MaRs are mainly produced by macrophages hence the derivation of the name from MAcrophage mediator RESolving INflammation. MaR1 exhibits potent anti-inflammatory, pro-resolving, analgesic and wound healing activities. Major cellular targets for the actions of MaR1 are vascular smooth muscle (VSM) cells and vascular endothelial cells. In these cells MaR1 attenuates the adhesion of monocytes to the endothelium induced by TNF-alpha. Maresin 1 also inhibits the production of reactive oxygen species by both VSM and endothelial cells. The major mechanism through which MaR1 exerts these effects is through down-regulation of the transcription factor, nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB). MaR2 has been shown to reduce neutrophil infiltration and to enhance macrophage-mediated phagocytosis of dead and dying cells, a process termed efferocytosis. Two related structures, the maresin-like mediators (MaR-L1 and MaR-L2), are generated when the maresins produced by macrophages are released and acted upon by leukocytes and platelets (Hong et al. 2014). These, together with 14,21-dihydroxy-DHAs, rescue the reparative function of diabetes-impaired macrophages in diabetic wound healing (Hong et al. 2014, Tian et al. 2011, Boniakowski et al. 2017)
Peroxisomes are small cellular organelles that are bounded by a single membrane and contain variable compositions of proteins depending on cell type. Peroxisomes function in oxidation of fatty acids, detoxification of glyoxylate, and synthesis of plasmalogens, glycerophospholipids containing an alcohol with a vinyl-ether bond (reviewed in Lohdi and Semenkovich 2014). All of the approximately 46 proteins contained in peroxisomal matrix are imported from the cytosol by a unique mechanism that does not require the imported proteins to be unfolded as they cross the membrane (Walton et al. 1995, reviewed in Ma et al. 2011, Fujiki et al. 2014, Baker et al. 2016, Dias et al 2016, Emmanoulidis et al. 2016, Erdmann 2016, Francisco et al. 2017). The incompletely characterized process appears to involve the transport of the proteins through a variably sized pore in the membrane comprising at least PEX5 and PEX14 (inferred from the yeast homologs in Meinecke et al. 2010, the yeast pore is reviewed in Meinecke et al. 2016). Oligomeric proteins are also observed to cross the peroxisomal membrane (Otera and Fujiki 2012) but their transport appears to be less efficient than monomeric proteins (Freitas et al. 2011, inferred from mouse homologs in Freitas et al. 2015, reviewed in Dias et al. 2016).In the cytosol, receptor proteins, PEX5 and PEX7, bind to specific sequence motifs in cargo proteins (Dodt et al. 1995, Wiemer et al. 1995, Braverman et al. 1997). The long and short isoforms of PEX5 (PEX5L and PEX5S) bind peroxisome targeting sequence 1 (PTS1, originally identified in firefly luciferase by Gould et al. 1989) found on most peroxisomal matrix proteins; PEX7 binds PTS2 (originally identified in rat 3-ketoacyl-CoA thiolase by Swinkels et al. 1991) found on 3 imported proteins thus far in humans. The long isoform of PEX5, PEX5L, then binds the PEX7:cargo protein complex (Braverman et al. 1998, Otera et al. 2000). PEX5S,L bound to a cargo protein or PEX5L bound to PEX7:cargo protein then interacts with a complex comprising PEX13, PEX14, PEX2, PEX10, and PEX12 at the peroxisomal membrane (Gould et al. 1996, Fransen et al. 1998, inferred from rat homologs in Reguenga et al. 2001).The ensuing step in which the cargo protein is translocated across the membrane is not completely understood. During translocation, PEX5 and PEX7 become inserted into the membrane (Wiemer et al. 1995, Dodt et al. 1995, Oliveira et al. 2003) and expose a portion of their polypeptide chains to the organellar matrix (Rodrigues et al. 2015). One current model envisages PEX5 as a plunger that inserts into a transmembrane barrel formed by PEX14, PEX13, PEX2, PEX10, and PEX12 (the Docking-Translocation Module) (Francisco et al. 2017).After delivering cargo to the matrix, PEX5 and PEX7 are recycled back to the cytosol by a process requiring mono-ubiquitination of PEX5 and ATP hydrolysis (Imanaka et al. 1987, Thoms and Erdmann 2006, Carvalho et al. 2007). PEX7 is not ubiquitinated but its recycling requires PEX5 mono-ubiquitination. A subcomplex of the Docking-Translocation Module comprising the RING-finger proteins PEX2, PEX10, and PEX12 conjugates a single ubiquitin to a cysteine residue of PEX5 (Carvalho et al. 2007, reviewed in Platta et al. 2016). The mono-ubiquitinated PEX5 and associated PEX7 are then extracted by the exportomer complex consisting of PEX1, PEX6, PEX26, and ZFAND6 (inferred from rat homologs in Miyata et al. 2012). PEX1 and PEX6 are members of the ATPases Associated with diverse cellular Activities (AAA) family, a group of proteins that use the energy of ATP hydrolysis to remodel molecular complexes. PEX1 and PEX6 form a hetero-hexameric ring, best described as a trimer of PEX1/PEX6 dimers (inferred from yeast in Platta et al. 2005, yeast homologs reviewed in Schwerter et al. 2017). Data on the yeast PEX1:PEX6 complex suggest that these ATPases use a substrate-threading mechanism to disrupt protein-protein interactions (Gardner et al. 2018). PEX7 is also then returned to the cytosol (Rodrigues et al. 2014). Once in the cytosol, ubiquitinated PEX5 is enzymatically deubiquitinated by USP9X and may also be non-enzymatically deubiquitinated by nucleophilic attack of the thioester bond between ubiquitin and the cysteine residue of PEX5 by small metabolites such as glutathione (Grou et al. 2012).Defects in peroxisomal import cause human diseases: Zellweger syndrome, neonatal adrenoleukodystrophy, infantile Refsum disease and rhizomelic chondrodysplasia punctata types 1 and 5 (Barøy et al. 2015, reviewed in Nagotu et al. 2012, Braverman et al. 2013, Wanders 2014, Fujiki 2016, Waterham et al. 2016)

 Statistics
Statistics